Genes & Cancer

Correlation between c-Met and ALDH1 contributes to the survival and tumor-sphere formation of ALDH1 positive breast cancer stem cells and predicts poor clinical outcome in breast cancer
Yuka Nozaki1,5, Shoma Tamori1,5, Masahiro Inada1, Reika Katayama1, Hiromi Nakane1, Osamu Minamishima1, Yuka Onodera1, Makoto Abe1, Shota Shiina1, Kei Tamura1, Daichi Kodama1, Keiko Sato3, Yasushi Hara4, Ryo Abe4, Ryoko Takasawa2, Atsushi Yoshimori6, Nariyoshi Shinomiya7, Sei-ichi Tanuma2 and Kazunori Akimoto1,5
1 Department of Medicinal and Life Science, Faculty of Pharmaceutical Sciences, Tokyo University of Science, Chiba, Japan
2 Department of Pharmacy, Faculty of Pharmaceutical Sciences, Tokyo University of Science, Chiba, Japan
3 Department of Information Sciences, Faculty of Science and Technology, Tokyo University of Science, Chiba, Japan
4 Research Institute for Biochemical Sciences, Tokyo University of Science, Chiba, Japan
5 Translational Research Center, Research Institute for Science& Technology, Tokyo University of Science, Chiba, Japan
6 Institute for Theoretical Medicine, Inc., Yokohama, Japan
7 Department of Integrative Physiology and Bio-Nano Medicine, National Defense Medical College, Saitama, Japan
Correspondence to: Kazunori Akimoto, email: [email protected]
Keywords: ALDH1, breast cancer, CSC, c-Met
Received: April 03, 2017
Accepted: August 13, 2017
Published: August 22, 2017
Copyright: Nozaki et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
ABSTRACT
c-Met is a receptor-type tyrosine kinase, which is involved in a wide range of cellular responses such as proliferation, motility, migration and invasion. It has been reported to be overexpressed in various cancers. However, the role of c-Met in breast cancer stem cells (CSCs) still remains unclear. We herein, show that c-Met expression is significantly elevated in Basal-like type of breast cancer in comparison with other subtypes. High expression of c-Met strongly correlated with the expression of two CSC markers, ALDH1A3 and CD133 in breast cancers. In addition, breast cancers at tumor stage III-IV expressing both c-Methigh and ALDH1A3high had poor prognosis. Furthermore, treatment with c-Met inhibitors (Crizotinib, Foretinib, PHA-665752 and Tivantinib) in MDA-MB157 cells with high c-Met protein expression resulted in significant suppression in cell viability, contrary to MDA-MB468 cells with low c-Met protein expression. These c-Met inhibitors also suppressed cell viability and tumor-sphere formation of ALDH1high breast cancer cells with high c-Met expression. These results suggest that c-Met in ALDH1 positive CSCs seems to play an important role in breast cancer repopulation. Therefore, we conclude that c-Met is a potential therapeutic target in ALDH1 positive breast CSCs.
INTRODUCTION
Breast cancer is one of the most common cancers occurring in women worldwide with 1.7 million new cases (25.2% of all cancers in women) and 0.5 million-cancer deaths (14.7% of all cancer death in women) according to an estimate from the International Agency for Research on Cancer (IARC) [1]. Breast cancer has been widely classified based on specific gene expression signature and receptor status. Based on PAM50 gene expression signature, breast cancer is categorized into six ‘’intrinsic’’ subtypes namely, Luminal A, Luminal B, HER2-enriched, Claudin-low, Basal-like, and Normal-like [2, 3, 4], of which, Basal-like type has poor prognosis [5]. Based on receptor status, breast cancer is categorized into estrogen receptor (ER)-positive type, progesterone receptor (PgR)-positive type, HER2 positive type, and triple-negative type (ER-negative, PgR- negative, HER2-negative) (TNBC). Among them, TNBC has the poorest prognosis. Notably, among 70-80% of Basal-like type of breast cancer has been reported to fall into TNBC category [6].
Tumors are comprised of population of cancer cells and distinct cancer stem cells (CSCs), which are largely undifferentiated tumorigenic cells with stem-like properties such as self-renewal and multipotency [7, 8]. Most CSCs are resistant to conventional anti-tumor treatments, chemo- and radio-therapies, which consequently leads tumor recurrence and metastasis. Therefore, the development of targeted therapies against CSCs is highly required to improve poor clinical outcome.
CSCs in breast tumor patients can be identified based on the expression of aldehyde dehydrogenase (ALDH) isoforms. ALDH1 has been reported to be enriched in CSCs of several cancer types, including breast cancer and is a potential CSC marker [9, 10, 11]. Among ALDH1 gene family, isoforms ALDH1A1 and ALDH1A3 are also known as CSCs markers in several cancers [11, 12, 13, 14]. Particularly, isoform ALDH1A3 has been reported to contribute significantly to ALDH1 activity in breast cancer cells and its expression significantly correlates with cancer type, tumor grade and metastasis in breast tumor patients [15]. On the other hand, there are controversial results regarding the involvement of ALDH1 in breast cancer subtypes [16, 17, 18].
c-Met is a receptor-type tyrosine kinase, which is involved in wide range of cellular responses such as proliferation, motility, migration, invasion and tumor angiogenesis [19, 20]. c-Met has been reported to be highly expressed and aberrantly activated in variety of cancers [21, 22, 23]. High expression of c-Met correlating with the expression of CSCs markers such as CD133, CD44, and ALDH1 has also been reported [24, 25, 26]. Furthermore, c-Met protein has been reported to be involved in biological processes of head and neck, and pancreatic CSCs [26, 27]. However, the relationship of c-Met with ALDH1 positive CSCs in breast cancer subtypes still remains unclear.
In this study, we show that high expression of c-Met correlates with the expression of ALDH1A3 in breast cancer. Patients with co-expression of c-Met and ALDH1A3 at tumor stage III-IV showed poor clinical outcome. Furthermore, c-Met inhibitors suppressed the viability and tumor-sphere formation of ALDH1high cells. These results suggest that c-Met is essential for the viability and tumor formation of ALDH1 positive breast CSCs. Therefore, c-Met protein is a promising therapeutic target for ALDH1 positive breast CSCs.
RESULTS
Correlation of c-Met with CSC markers at gene expression level in human breast cancers
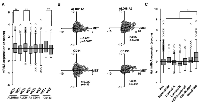
To investigate the association of c-Met with CSC markers such as ALDH1A1, ALDH1A3, CD44, and CD133 at gene expression levels in human breast cancers, we analyzed mRNA data and the clinical information of 1904 patients of breast cancers from cBioPortal for Cancer Genomics [28, 29]. As shown in Figure 1A, high expression of c-Met (MET+) correlated with expression of CSC markers, ALDH1A1 (p < 0.001), ALDH1A3 (p < 0.001), and CD133 (p < 0.001) in breast cancers. In addition, scatter plots analysis also indicated that c-Met expression correlated with ALDH1A1 (p = 0.0077), ALDH1A3 (p < 0.001) and CD133 expression (p < 0.001) (Figure 1B and Table 1). c-Met expression was also found to be associated with several undifferentiated markers, such as KLF4, c-Myc, Notch1, Notch3, and BMI1 (Table1). Next we examined the mRNA expression level of c-Met in the specific breast cancer subtypes. As shown in Figure 1C, c-Met mRNA was found to be enriched in Basal-like type in comparison with other subtypes, such as Normal-like, Luminal A, Luminal B, HER2-enriched, and Claudin-low.
Correlation of c-Met with ALDH1A3 and CD133 at gene expression level in breast cancer at tumor stage III-IV
Since overexpression of c-Met contributes to cancerous progression [21, 22, 23], we next examined c-Met expression at various tumor stages. Among early tumor stage lesions (0, I, II; n = 1279), 45% were c-Met+(n = 573), contrary to 59% of c-Met+ (n = 74) at tumor late stage lesions (III, IV; n = 124). As c-Met+ tumor lesions were higher in tumor stage III-IV, in contrast with stage 0, I, and II, we next focused to analyze the relationship between c-Met gene expression and CSC markers in breast cancer subtypes at tumor stage III-IV. c-Met mRNA was found to be enriched in Basal-like type in comparison with other subtypes at stage III and IV (Figure 2A). As shown in Figure 2B, c-Met+ strongly correlated with ALDH1A3+(p < 0.001). c-Met+ also weakly associated with CD133+ (p = 0.0025). Scatter plots analysis also indicated that c-Met expression correlated with ALDH1A3 (p < 0.001) and CD133 expression (p < 0.001) (Figure 2C and Table 1). c-Met expression was also found to be associated with several undifferentiated markers, such as Notch1, Oct4, and BMI1 (Table 1). These results indicate that c-Met plays important roles in ALDH1 and/or CD133 positive CSCs.
Co-expression of c-Methigh and ALDH1A3high indicated poor prognosis
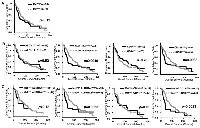
Further, we next performed Kaplan-Meier analysis of c-Met and CSC markers at tumor stage III-IV. c-Methigh patients did not show poor prognosis (p = 0.11) (Figure 3A), whereas ALDH1A3high(p = 0.0049) and CD133high(p = 0.0088) showed poor prognosis (Figure 3B). Interestingly, co-expression of both c-Methigh with ALDH1A3high (p = 0.0065), and with CD133high (p = 0.0023) indicated poor prognosis (Figure 3C). These results indicate that c-Met plays important roles in cancerous progression and contributed to the poor prognosis in ALDH1 positive and/or CD133 positive breast CSCs. Since the role of c-Met in biological properties of CD133 positive CSCs is reported [24, 32, 33], hence, we focused on investigating the roles of c-Met in ALDH1 positive breast CSCs.
c-Met inhibitors suppressed viability of ALDH1 positive CSCs
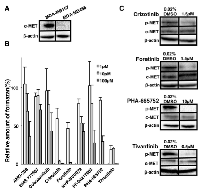
To reveal the role of c-Met in CSCs, we used MDA-MB157 and MDA-MB468 cell lines derived from human Basal-like type of breast cancer. c-Met protein was found to be highly expressed in MDA-MB157 cells in contrast to MDA-MB468 cells (Figure 4A). Next, we examined the effects of nine c-Met inhibitors on the viability of MDA-MB157 cells expressing higher c-Met protein (Table 2). Four c-Met inhibitors such as Crizotinib, Foretinib, PHA-665752 and Tivantinib strongly suppressed the viability of MDA-MB157 cells (Figure 4B). These results were consistent with the results of inhibition of c-Met phosphorylation level (indicating its activity) on treatment with c-Met inhibitors in MDA-MB157 cells (Figure 4C). Therefore, we next examined the inhibitory effects of these four c-Met inhibitors on the viability of ALDH1high cells derived from MDA-MB157 and MDA-MB468 cell lines. Isolated ALDH1high cells derived from both MDA-MB157 and MDA-MB468 cell lines showed CSCs properties such as self-renewal, multi-differentiation, and tumorigenesis (Supplementary Figure2A and 2B) as previously reported [9]. Interestingly, both c-Met and p-Met expression is higher in ALDH1high cells than ALDH1low cells (Figure 5A). The result suggests that ALDH1high cells have high activity of c-Met. The c-Met inhibitors except for Tivantinib suppressed viability of ALDH1high cells in both cell lines. The 50% cell growth inhibitory concentrations (IC50) of Crizotinib, Foretinib, and PHA-665752 were found to be lower in MDA-MB157 cells expressing higher c-Met protein than that in MDA-MB468 cells expressing lower c-Met protein (Figure 5B-D). Interestingly, Tivantinib specifically suppressed the viability of ALDH1high MDA-MB157 cells. These results suggest that c-Met is necessary for the viability of ALDH1 positive breast CSCs.
c-Met inhibitors suppressed tumor-sphere formation of ALDH1 positive CSCs
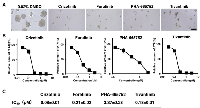
To investigate the role of c-Met in tumor formation of ALDH1 positive CSCs, we next examined the inhibitory effects of aforementioned inhibitory compounds on tumor-sphere formation in ALDH1 positive CSCs derived from MDA-MB157 in vitro system. As shown in Figure 6A and 6B, the inhibitory compounds were observed to suppress tumor-sphere formation. The IC50 values of these compounds for tumor-sphere formation were 0.18 μM (Crizotinib), 0.21 μM (Foretinib), 3.4μM (PHA-665752), and 0.18 μM (Tivantinib) (Figure 6C). These results suggest that c-Met is essential for tumor-sphere formation of ALDH1 positive CSCs in breast cancer cells.
Taken together with aforementioned results, it can be inferred that c-Met is specifically essential for cell viability and tumor-sphere formation of ALDH1 positive human breast CSCs.
DISCUSSION
High expression of c-Met correlated with the expression of ALDH1A3 in Basal-like type of breast cancer (Figure 1C). Since breast cancer stem cells exhibit a Basal-like phenotype [34], our result may thus provide new insights into the role of c-Met in ALDH1 positive CSCs of Basal-like type of breast cancer. It has been reported that knock-down of c-Met by siRNA and inhibitor treatment results in decrease of ALDH1A3 gene expression and ALDEFLUOR activity in pancreatic cancer cell lines with high levels of c-Met [25]. Similarly, high c-Met expression and its activation are also suggested to be involved in the promotion of ALDH1A3 gene expression in Basal-like type of breast cancer.
Several studies have reported that patients with higher expression of ALDH1 have poor prognosis in several cancers [9, 35]. In our study, Kaplan-Meier analysis revealed that patients with high ALDH1A3 expression at tumor stage III-IV had poor outcome (p = 0.0049, Figure 3B). Similarly, patients expressing both c-Met and ALDH1A3 at tumor stage III-IV had poor prognosis (p = 0.0065, Figure 3C). c-Met was found to be enriched in Basal-like type in comparison with other subtypes (Figure 1 and 2). However, patients expressing high c-Met and ALDH1A3 in Basal-like type did not show poor outcome (p = 0.20; n = 199, Supplementary Figure 1D), which could be attributed to no correlation between c-Met and ALDH1A3 in Basal-like type (Supplementary Figure 1A, p = 0.58, r = 0.039). In spite of no correlation between c-Met and ALDH1A3 expression, major population of Basal-like type patients expressed high expression of c-Met and ALDH1A3 (n = 93 in 199). Therefore, c-Met may play an important role in cancerous progression in Basal-like type. On the other hands, at tumor stage III-IV, of total analyzed patient samples (n = 124), each subtypes were distributed as follows; Luminal A (23%, n = 28), Luminal B (28%, n = 35), HER2-enriched (17%, n = 21), Claudin-low (16%, n = 20), Normal-like (5%, n = 6), and Basal-like (11%, n = 14). Therefore, co-expression of both c-Met and ALDH1A3 at late tumor stages may contribute to poor clinical outcome not only in Basal-like but also in other subtypes. Since efficacy of chemotherapy at cancer spreading stage III-IV is extremely crucial, targeting c-Met in ALDH1 positive breast CSC may possibly decrease the severity of metastatic breast cancer and hence may lead to the survival of breast cancer patients. In addition, previous studies reported that ALDH1 is required for maintaining a drug-resistant cell subpopulation of stomach and breast cancer cells [36, 37, 38, 39]. Therefore, considering this, the drug resistance characteristics of breast cancers expressing c-Met and ALDH1A3 should be analyzed in detail in the future for targeted cancer therapy.
We found that c-Met inhibitors suppressed cell viability and tumor-sphere formation of ALDH1high cells (Figure 5 and 6). ALDH1 enzyme catalyzes the oxidation of aldehydes into corresponding acetic acids, and is involved in detoxification of toxic aldehyde intermediates produced in cancer cells. Recent studies reported that ALDH1 decreases ROS levels in various cancer cells and metabolizes toxic aldehydes formed by lipid peroxidation generated from intracellular lipids due to ROS [36, 40]. Since we observed strong correlation between ALDH1 and c-Met, use of c-Met inhibitors in ALDH1high cells may have accumulated ROS and toxic aldehydes, which consequently may have lead to the induction of apoptosis in cancer cells.
Thus, it is suggested that c-Met plays an important role in ALDH1 positive breast CSCs. Although the ALDH1high cells derived from MDA-MB157 and MDA-MB468 cells have been cultured in vitro in the presence of FBS, no loss in CSCs properties was observed (Supplementary Figure2A and 2B). Since, loss of stem cell property due to long term culture of cells in in vitro in the presence of FBS has been reported, hence appropriate measures should be taken for long term culture of CSCs.
Among c-Met inhibitors, Crizotinib, Foretinib, PHA-665752 and Tivantinib, only Tivantinib specifically suppressed viability of high c-Met expressing MDA-MB157 cells as compared to low c-Met expressing MDA-MB468 cells (Figure 5). These results may depend on the inhibitory mechanisms of Tivantinib against c-Met activity. The c-Met inhibitors except Tivantinib are ATP competitor that docks to active site of c-Met kinase. ATP competitors generally inhibit the activity of other kinases and function of ATP associated molecules. In fact, Crizotinib, Foretinib and PHA-665752 strongly suppress the cell viability by inhibition of other kinases and ATP associated molecules (Table 2). On the other hand, Tivantinib, a non-ATP competitor, inhibits c-Met autophosphorylation and is highly selective for the inactive or non-phosphorylated form of c-Met by binding to ATP-binding cleft [41, 42]. Furthermore, the specific inhibitory effect of Tivantinib is profiled against 230 human kinases [41]. Non-ATP competitor such as Tivantinib binding to allosteric site must be explored further, as it may contribute to develop specific drugs targeting to c-Met in the future.
CONCLUSION
In this study, we showed that high expression of c-Met correlated with the expression of ALDH1A3 in Basal-like type of breast cancer. Patients with co-expression of c-Met and ALDH1A3 at tumor stage III-IV showed poor clinical outcome. Furthermore, c-Met inhibitors suppressed the cell viability and tumor-sphere formation of ALDH1high cells. These results suggest that c-Met is essential for the viability and tumor formation of ALDH1 positive CSCs. Therefore, c-Met protein is potential therapeutic target for ALDH1 positive breast CSCs.
MATERIALS AND METHODS
Cell culture
Human Basal-like type of breast cancer cell lines (MDA-MB157 and MDA-MB468) were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). Cell lines were grown in Dulbecco’s Modified Eagle Medium (DMEM) medium supplemented with 10% fetal bovine serum (FBS) (Biosera, Dominican Republic) and penicillin/streptomycin. Cells were cultured at 37°C in a humidified atmosphere with 95% air/5% CO2.
c-Met inhibitors and antibodies
c-Met inhibitors (AMG-208, BMS 777607, Cabozantinib, Crizotinib, Foretinib, NVP-BVU972, PF-04217903, PHA-665752, Tivantinib) were purchased from Namiki Inc. (Japan). All compounds dissolved in DMSO. Rabbit polyclonal c-Met antibody was purchased from Santa Cruz Inc. (USA). Rabbit monoclonal phospho-Met (Tyr1234/1235) antibody, HRP-conjugated anti-rabbit IgG and anti-mouse IgG were purchased from Cell Signaling Technology (USA). Mouse monoclonal β-actin antibody was obtained from Wako Inc. (Japan).
Flow cytometry
Cells were exfoliated from culture dish by accutase (Innovative Cell Technology) and filtered through 40µm cell strainers (Greiner) to obtain single cells. The ALDH1high cells were isolated from MDA-MB157 and MDA-MB468 cells by ALDEFLUOR assay kit (Stem Cell Technology) or AldeRed ALDH detection assay kit (MERCK) according to the manufacturer’s instructions. Briefly, cells (2×106) were incubated with the substrate for ALDH1 (5μL substrate/mL medium) for 30 min at 37° C. As a negative control for the ALDEFLUOR assay and AldeRed assay, cells were incubated with ALDH1 inhibitor, diethylaminobenzaldehyde (DEAB). The ALDH1high cells were sorted by cell sorter (FACS AriaII, BD Bioscience) by taking the negative control into consideration. The analysis of CD10/EpCAM positive cells from MDA-MB157 and MDA-MB468 cells. Suspended MDA-MB468 cells (1×106) were incubated with anti-CD10 (APC) (BD Bioscience) and anti-EpCAM (PE) (BD Bioscience) for 1hr on ice, after which the sample was washed with fresh FACS buffer (2%FBS in 1×PBS (-)). For this experiments, cells were analyzed using a FACS Calibur (BD Bioscience).
WST-8 assay
Cells (3×105/well) were seeded into 96 well culture plate (Sigma). One day post seeding, cells were treated with c-Met inhibitors for 3, 5, and 7 days. Cell viability was detected by WST-8 assay (Cell Counting Kit-8 (DOJINDO)). The formazan dye formed was measured by ARVOTM MX (PerkinElmer) at 450 nm. Numerical values of test groups are shown with respect to 0.02% DMSO treated group.
Immunoblotting
Cells were dissolved in RIPA buffer (50 mM Tris (pH 8.0), 150 mM NaCl, 0.5 w/v% sodium deoxycholate, 0.1 w/v % SDS, 1.0 w/v % Nonidet P-40 and protease inhibitor cocktail (Thermo Fisher)). Eight μg of whole cell lysate proteins was electrophoresed by SDS-PAGE (8% gel) and transferred to Immobilon-P Transfer Membrane (Millipore) or Immobilon-FL Transfer Membrane (Millipore). The transferred membranes were then blocked with 5% BSA in TTBS (25 mM Tris (pH 7.5), 140 mM NaCl, 2.5 mM KCl and 0.1% Tween 20) and incubated with the primary antibodies. The membranes were then probed with the horseradish peroxidase-conjugated secondary antibody. Specific signals were detected by chemiluminescence reagent, such as Immunostar LD/ Immunostar Basic (Wako) using ChemiDoc MP (Bio-Rad).
Tumor-sphere culture
Tumor-spheres were grown in DMEM culture medium containing 10% FBS, penicillin and streptomycin, 0.6% methyl cellulose (Wako), and 0.05 mM 2-mercaptoethanol (Sigma) at 37°C in a humidified atmosphere with 95% air/5% CO2. ALDH1high cells (1x103/well) were seeded and cultured in ultra low attachment 96-well plate (Greiner) for 6 days with or without inhibitory compounds. CellTiter-Glo® luminescence assay (Promega) was performed by TR717 Micro plate Luminometer (TROPIX) using 96 well Micro-assay-plate (Greiner). Numerical values of test groups are shown with respect to 0.02% DMSO treated group.
Analysis of gene expression data
Gene expression data was analyzed using METABRIC, Nature 2012 & Nat. Commun. 2016 dataset deposited in cBioPortal [28, 29, 30, 31]. Clinical data of the breast cancer patients used in our present study are summarized in Table S1. The median age at diagnosis was 61.1 years (aged 21.9 to 96.3 years). The dataset contains mRNA expression data of 1,904 primary breast tumor samples (patients) with details of breast cancer subtype (Normal-like, n = 140; Luminal A, n = 679; Luminal B, n = 461; HER2-enriched, n = 220; Claudin-low, n = 199; Basal-like, n = 199; Not classified, n = 6). We retrieved the mRNA expression (Z-scores) of genes and evaluated co-expression of c-Met and several stem cell markers in either all or each of the tumor stage groups. We defined the c-Met expression as follows; all stage patients were divided into c-Met+(c-Met mRNA expression Z-score>0, n = 837) and c-Met-(c-Met mRNA expression Z-score < 0, n = 1067) in Figure 1A, 1C and Table1. Tumor stage III and IV patients were classified into c-Met+(c-Met mRNA expression Z-score>0, n = 74) and c-Met-(c-Met mRNA expression Z-score < 0, n = 50) in Figure 2A, 2B and Table1. Pearson’s correlation coefficiency was calculated for these expression levels for the subtypes in Figure 1B, 2C and Table1. We also compared c-Met expression in all or stage III-IV groups. Quantitative variables were analyzed by Tukey’s test. Data with p value less than 0.05 were considered significant. Survival curves were plotted by the Kaplan-Meier method and compared by the Gehan-Breslow generalized Wilcoxon test using BellCurve for Excel ver2.11. “High” and “low” were defined as the upper top 40% and the lower 60% of Z-score respectively, in several genes at stages III-IV breast cancer patients. Follow-up period after diagnosis ranged from 5.8 to 274.3 months stages III-IV breast cancer patients.
ACKNOWLEDGMENTS
We would like to thank Dr. T. Sato for helpful discussion on statistical analysis. We are also extremely grateful to Dr. B. Shashni for proof reading the article. This work was supported by the MEXT-Supported Program for the Strategic Research Foundation at Private Universities, 2014-2018. This work was supported in part by Grant-in-Aid for Scientific Research, Japan Society for the Promotion of Science to N.S. (#23300365, #15H04315).
CONFLICTS OF INTEREST
There is no conflict of interest.

- 1. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015; 136: E359-386. [PubMed]
- 2. Supervised Risk Predictor of Breast Cancer Based on Intrinsic Subtypes. J Clin Oncol. 2009; 27: 1160–1167. [PubMed] https://doi.org/10.1200/JCO.2008.18.1370.
- 3. Clinical implications of the intrinsic molecular subtypes of breast cancer. Breast. 2015; 24: S26–S35. [PubMed]
- 4. Intrinsic Subtypes from PAM50 Gene Expression Assay in a Population-Based Breast Cancer Cohort: Differences by Age, Race, and Tumor Characteristics. Cancer Epidemiol Biomarkers Prev. 2015; 23: 714-724. [PubMed] https://doi.org/10.1158/1055-9965.EPI-13-1023.
- 5. Genomics in breast cancer-therapeutic implications. Nature Reviews Clinical Oncology. 2005; 2: 26-33. [PubMed]
- 6. Basallike and triple-negative breast cancers: a critical review with an emphasis on the implications for pathologists and oncologists. Mod Pathol. 2011; 24: 157-167. [PubMed]
- 7. Cancer Stem Cells: Current Status and Evolving Complexities. Cell Stem Cell. 2012; 10: 717-728. [PubMed]
- 8. Stem cells, cancer, and cancer stem cells. Nature. 2001; 1: 105-111. [PubMed]
- 9. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell. 2007; 1: 555–567. [PubMed] https://doi.org/10.1016/j.stem.2007.08.014.
- 10. Aldehyde dehydrogenase 1 is a tumor stem cell-associated marker in lung cancer. Mol Cancer Res. 2009; 7: 330-338 [PubMed] https://doi.org/10.1158/1541-7786.MCR-08-0393.
- 11. ALDH1A1 Positive Cell Population Is Enriched in Tumorinitiating Cells and Associated with Progression of Bladder Cancer. Cancer Epidemiol Biomarkers Prev. 2010; 19: 327-337. [PubMed] https://doi.org/10.1158/1055-9965.EPI-09-0865.
- 12. Targeting Aldehyde Dehydrogenase Cancer Stem Cells in Ovarian Cancer. Molecular Cancer Therapeutics. 2010; 9: 3186-3199. [PubMed] https://doi.org/10.1158/1535-7163.MCT-10-0563.
- 13. Aldehyde Dehydrogenase Is Regulated by β-Catenin/TCF and Promotes Radioresistance in Prostate Cancer Progenitor Cells. Cancer Res. 2015; 75: 1482-1494. [PubMed]
- 14. Aldehyde dehydrogenase 1A3 influences breast cancer progression via differential retinoic acid signaling. Mol Oncol. 2015; 9: 17-31. [PubMed] https://doi.org/10.1016/j.molonc.2014.07.010.
- 15. Aldehyde Dehydrogenase Activity of Breast Cancer Stem Cells is Primarily Due to Isoform ALDH1A3 and Its Expression is Predictive of Metastasis. Stem Cells. 2011; 29: 32-45. [PubMed]
- 16. Breast Cancer Stem Cells and Intrinsic Subtypes: Controversies Rage On. Current Stem Cell Research & Therapy. 2009; 4: 50-60. [PubMed]
- 17. Cancer stem cell markers are associated with adverse biomarker profiles and molecular subtypes of breast cancer. Breast Cancer Research and Treatment. 2012; 136: 407-417. [PubMed]
- 18. Breast cancer stem cell markers CD44, CD24 and ALDH1: expression distribution within intrinsic molecular subtype. J Clin Pathol. 2011; 64: 937-946. [PubMed]
- 19. An overview of the c-MET signaling pathway. Therapeutic Advances in Medical Oncolog. 2011; 3: 7-19. [PubMed] https://doi.org/10.1177/1758834011422556.
- 20. Scatter factor/hepatocyte growth factor in brain tumor growth and angiogenesis. NeuroOncology. 2005; 7: 436-451. [PubMed] https://doi.org/10.1215/S1152851705000050.
- 21. Genomic amplification of MET with boundaries within fragile site FRA7G and upregulation of MET pathways in esophageal adenocarcinoma. Oncogene. 2006; 25: 409-418. [PubMed]
- 22. Met gene copy number predicts the prognosis for completely resected non-small cell lung cancer. Cancer Sci. 2008; 99: 22802285. [PubMed]
- 23. Developing c-MET pathway inhibitors for cancer therapy: progress and challenges. Trends Mol Med. 2010; 16: 37-45. [PubMed]
- 24. In Vivo c-Met Pathway Inhibition Depletes Human Glioma Xenografts of Tumor-Propagating Stem-Like Cells. Transl Oncol. 2013; 6: 104-111. [PubMed]
- 25. Fibulin-3 negatively regulates ALDH1 via c-MET suppression and increases γ-radiation-induced sensitivity in some pancreatic cancer cell lines. Biochem Biophys Res Commun. 2014; 454: 369-375. [PubMed]
- 26. c-Met pathway promotes self-renewal and tumorigenecity of head and neck squamous cell carcinoma stem-like cell. Oral Oncology. 2014; 50: 633639. [PubMed]
- 27. Susceptibility of pancreatic cancer stem cells to reprogramming. Cancer Sci. 2015; 106: 1182–1187. [PubMed] https://doi.org/10.1111/cas.12734.
- 28. The genomic and transcriptomic architecture of 2,000 breast tumours reveals novel subgroups. Nature. 2012; 486: 346–352. [PubMed] https://doi.org/10.1038/nature10983.
- 29. The somatic mutation profiles of 2,433 breast cancers refine their genomic and transcriptomic landscapes. Nat Commun. 2016; 7: 11479. [PubMed] https://doi.org/10.1038/ncomms11908.
- 30. The cBio Cancer Genomics Portal: an open platform for exploring multidimensional cancer genomics data. Cancer Dicov. 2012; 2: 401-404. [PubMed] https://doi.org/10.1158/2159-8290.CD-12-0095.
- 31. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal. 2013; 6: 11. [PubMed] https://doi.org/10.1126/scisignal.2004088.
- 32. Combination of dasatinib and curcumin eliminates chemo-resistant colon cancer cells. J Mol Signal. 2011; 20: 6:7 [PubMed] https://doi.org/10.1186/1750-2187-6-7.
- 33. c-MET protects breast cancer cells from apoptosis induced by sodium butyrate. PLoS One. 2012; 7: e30143. [PubMed] https://doi.org/10.1371/journal.pone.0030143.
- 34. Farrar. Cancer Stem Cell. Cambridge University press. 2010; 50-51.
- 36. The Cancer Stem Cell Marker Aldehyde Dehydrogenase Is Required to Maintain a Drug-Tolerant Tumor Cell. Cancer Res. 2014; 74: 3579-3590. [PubMed]
- 37. Aldehyde dehydrogenase high gastric cancer stem cells are resistant to chemotherapy. Int J Oncol. 2013; 42: 1437-1442. [PubMed]
- 38. Inhibition of aldehyde dehydrogenase (ALDH) activity reduces chemotherapy and radiation resistance of stem-like ALDHhiCD44+ human breast cancer cells. Breast Cancer Res Treat. 2012; 133: 75-87. [PubMed]
- 39. Association of Breast Cancer Stem Cells Identified by Aldehyde Dehydrogenase 1 Expression with Resistance to Sequential Paclitaxel and EpirubicinBased Chemotherapy for Breast Cancers. Clin Cancer Res. 2009; 15: 4234-4241. [PubMed]
- 40. Aldehyde Dehydrogenases in Cellular Responses to Oxidative/ electrophilic Stress. Free Radic Biol Med. 2013; 56: 89101. [PubMed] https://doi.org/10.1016/j.freeradbiomed.2012.11.010.
- 41. ARQ 197, a Novel and Selective Inhibitor of the Human c-Met Receptor Tyrosine Kinase with Antitumor Activity. Mol Cancer Ther. 2010; 9: 1544-1553. [PubMed]
- 42. Discovery of a novel mode of protein kinase inhibition characterized by the mechanism of inhibition of human mesenchymal-epithelial transition factor (c-Met) protein autophosphorylation by ARQ197. J. Biol. Chem. 2011; 286: 20666-20676. [PubMed] https://doi.org/10.1074/jbc.M110.213801.
- 43. A drug resistance screen using a selective MET inhibitor reveals a spectrum of mutations that partially overlap with activating mutations found in cancer patients. Cancer Res. 2011; 71: 5255-5264. [PubMed]
- 44. Discovery of N-(4-(2-amino3-chloropyridin-4-yloxy)-3-fluorophenyl)-4-ethoxy-1-(4fluorophenyl)-2-oxo-1,2-dihydropyridine-3-carboxamide (BMS-777607), a selective and orally efficacious inhibitor of the Met kinase superfamily. J Med Chem. 2009; 52: 1251-1254. [PubMed]
- 45. Albrecht, Jean-Christophe Harmange, David Bauer, Loren Berry, Christiane Bode, Alessandro A. Boezio, April Chen, Deborah Choquette, Isabelle Dussault, Cary Fridrich, Satoko Hirai, Doug Hoffman, Jay F. Larrow, et al. Discovery and Optimization of Triazolopyridazines as Potent and Selective Inhibitors of the c-Met Kinase. J. Med. Chem. 2008; 51: 2879–2882 [PubMed]
- 46. Cabozantinib overcomes crizotinib resistance in ROS1 fusion positive cancer. Clin Cancer Res. 2015; 21: 166-174. [PubMed] https://doi.org/10.1158/1078-0432.CCR-14-1385.
- 47. Novel mechanism of Lapatinib resistance in HER2-positive breast tumor cells: activation of AXL. Cancer Res. 2009; 69: 6871-6878. [PubMed] Novel mechanism of Lapatinib resistance in HER2-positive breast tumor cells: activation of AXL
- 48. Inhibition of tumor cell growth, invasion, and metastasis by EXEL-2880 (XL880, GSK1363089), a novel inhibitor of HGF and VEGF receptor tyrosine kinases. Cancer Res. 2009; 69: 8009-8016. [PubMed]
- 49. Discovery of a novel class of exquisitely selective mesenchymal-epithelial transition factor (c-MET) protein kinase inhibitors and identification of the clinical candidate 2-(4-(1-(quinolin-6-ylmethyl)-1H-[1, 2, 3]triazolo[4,5-b]pyrazin-6-yl)-1H-pyrazol-1-yl)ethanol (PF04217903) for the treatment of cancer. J Med Chem. 2012; 55: 8091-8109. [PubMed]
- 50. Sensitivity of Selected Human Tumor Models to PF-04217903, a Novel Selective c-Met Kinase Inhibitor. Mol Cancer Ther. 2012; 11: 1036-1047. [PubMed]
- 51. An orally available small-molecule inhibitor of c-Met, PF-2341066, exhibits cytoreductive antitumor efficacy through antiproliferative and antiangiogenic mechanisms. Cancer Res. 2007; 67: 4408-4417. [PubMed]
- 52. Cytoreductive antitumor activity of PF-2341066, a novel inhibitor of anaplastic lymphoma kinase and c-Met, in experimental models of anaplastic large-cell lymphoma. Mol Cancer Ther. 2007; 6: 3314-3322. [PubMed]
- 53. A Selective Small Molecule Inhibitor of c-Met Kinase Inhibits c-Met-Dependent Phenotypes in Vitro and Exhibits Cytoreductive Antitumor Activity in Vivo. Cancer Res. 2003; 63: 7345-7355. [PubMed]
- 54. Amplification of MET may identify a subset of cancers with extreme sensitivity to the selective tyrosine kinase inhibitor PHA665752. PNAS. 2006; 103: 2316-2321. [PubMed] https://doi.org/10.1073/pnas.0508776103.
Last Modified: 2017-09-27 20:30:48 EDT
 All site content, except where otherwise noted, is licensed under a Creative Commons Attribution 4.0 License.
All site content, except where otherwise noted, is licensed under a Creative Commons Attribution 4.0 License.